Gamle øyne, moderne design: Trilobittenes bemerkelsesverdige syn
Emily Reeves 23. oktober 2025 Oversatt herfra.

Trilobitter, utdødde marine dyr som først dukker opp i de kambriske berglagene, gled over gamle havbunner. Vanligvis bevarer fossilisering bare deres harde eksoskjeletter, og etterlater lite spor etter delikate strukturer som sanseorganer. Imidlertid avslørte en usedvanlig godt bevart Aulacopleura koninckii, en trilobittart, et sammensatt øye med slående likheter med moderne insekter og krepsdyr.
I en studie publisert i Nature -lenke, beskrev hovedforfatter Brigitte Schoenemann og teamet hennes denne trilobittens øye som et "fullstendig moderne" visuelt system, sammenlignbart med levende bier, øyenstikkere og dagaktive krepsdyr. De sier:
"Vi viser at denne paleozoiske trilobitten i prinsippet var utstyrt med en fullstendig moderne type visuelt system, et sammensatt øye som kan sammenlignes med levende bier, øyenstikkere og mange dagaktive krepsdyr. Det er et eksempel på utmerket bevaring, og vi håper at dette manuskriptet vil være et utgangspunkt for mer forskningsarbeid på fossile bevis, og for å utvikle en dypere forståelse av synets utvikling."
Bilde 1. Trilobitt øyne
Et svært moderne øye
 Fossilets utsøkte bevaring gjorde det mulig for forskere å identifisere trekk ved et apposisjons-sammensatt øye, en type som finnes hos moderne leddyr som reker, hummer og sirisser. Det apposisjons-sammensatte øyet fungerer ved å dele synsfeltet inn i tusenvis av små segmenter, som hvert behandles av et ommatidium (ommatidium er en individuell strukturell enhet som utgjør det sammensatte øyet til et leddyr), for å skape et bredt, bevegelsesfølsomt mosaikkbilde. Selv om sammensatte øyne ikke forstås å ha like høy oppløsning som menneskeøyne, gir disse øynene et nesten 360-graders panoramabilde, raskere behandling av visuelle endringer og evnen til å oppdage polarisert og ultrafiolett lys hos noen arter. Disse egenskapene gjør det mulig for leddyr å oppfatte mønstre og navigere på måter utenfor menneskelig syn.
Fossilets utsøkte bevaring gjorde det mulig for forskere å identifisere trekk ved et apposisjons-sammensatt øye, en type som finnes hos moderne leddyr som reker, hummer og sirisser. Det apposisjons-sammensatte øyet fungerer ved å dele synsfeltet inn i tusenvis av små segmenter, som hvert behandles av et ommatidium (ommatidium er en individuell strukturell enhet som utgjør det sammensatte øyet til et leddyr), for å skape et bredt, bevegelsesfølsomt mosaikkbilde. Selv om sammensatte øyne ikke forstås å ha like høy oppløsning som menneskeøyne, gir disse øynene et nesten 360-graders panoramabilde, raskere behandling av visuelle endringer og evnen til å oppdage polarisert og ultrafiolett lys hos noen arter. Disse egenskapene gjør det mulig for leddyr å oppfatte mønstre og navigere på måter utenfor menneskelig syn.
Bilde 2. Trilobitt-avtrykk fra Kambrium
Ved undersøkelse av A. koninckii-øyet, som hadde bevart bløtvevsstrukturer, oppdaget forskerne et svært moderne sammensatt øye. Antyder denne oppdagelsen at prinsippene for sammensatt syn dukket opp for nesten en halv milliard år siden, slik forfatterne konkluderte? Det ville antyde at slike komplekse visuelle systemer utviklet seg innen korte evolusjonære tidsrammer i løpet av den kambriske perioden -lenke . Denne raske fremveksten utfordrer dagens forståelse, ettersom bevis for den kreative evnen til tilfeldig mutasjon og naturlig utvalg til å produsere slike intrikate strukturer i løpet av et kort vindu fortsatt er begrenset og omdiskutert. Funnet av slik moderne synsutstyr i en gammel organisme utfordrer ytterligere den vanlige evolusjonære historien om synets opprinnelse.
For mer om den kambriske eksplosjonen og dens implikasjoner, vurder å lese fra Darwins tvil -her.
Optimalisering: Den tekniske forklaringen på "evolusjon som skjer foran øynene våre".
Av Emily Reeves 22. oktober 2025. Oversatt herfra. {kursiv og understreking ved oversetter.}
Du kan ikke være god i alt. Det finnes avveininger. For eksempel har en racerbil fordeler på en racerbane, men en jeep har fordeler på en gjørmete, steinete vei. Akkurat som en bil kan designes for fart på en racerbane eller for terrengkjøring, finnes det designavveininger i biologi. Å forstå disse avveiningene og deres optimalisering for populasjoner er nøkkelen til å forstå eksempler, ofte rapportert av vitenskapelige medier, på "evolusjon som skjer foran øynene våre".
Viktige forbindelser mellom avveininger og optimalitet
Avveininger er nødvendige i multi-optimalitet. Med andre ord må et system optimaliseres over flere variabler, noe som innebærer avveininger mellom separate mål. Avveininger er fundamentalt knyttet til optimalitet fordi de definerer begrensningene som et system må operere innenfor, for å oppnå best mulig ytelse. I både ingeniørfag og biologi betyr det å oppnå en optimal løsning ofte å balansere konkurrerende mål, i stedet for å maksimere eller minimere en enkelt variabel som hastighet.
Om et design er optimalt, avhenger da av oppgavene som skal utføres. Tenk igjen på bileksemplet. Hvis du plasserer en racerbil på en steinete, svingete vei opp til en gammel gruveby, vil den ha svært lav "egnethet". Sett en jeep på samme vei, og den vil ha høy "egnethet", fordi designet er tilpasset oppgavene som skal utføres. Dermed er optimalitet avhengig av miljøet objektet eller organismen plasseres i. Eller jeg kan si at optimalitet er avhengig av målene som designeren optimaliserer den for.
Bilde 3. Arketyper og Pareto-optimalitet
En introduksjon til ytelsesrom og pareto-optimalitet
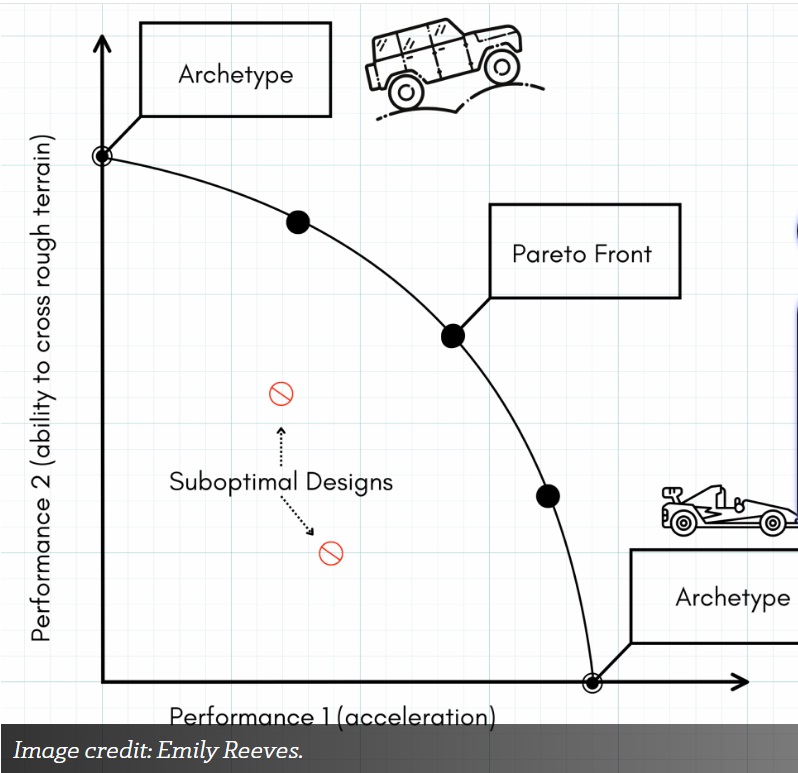 Jeg baserer mye av den følgende diskusjonen på systembiolog Uri Alons arbeid (Alon 2019; Shoval et al. 2012). La oss forestille oss at vi designer biler: en jeep og en racerbil. De to ytelsesegenskapene vi er interessert i er akselerasjon og evnen til å krysse ulendt terreng. Hvis vi plotter ytelse 1 på x-aksen og ytelse 2 på y-aksen, kan vi kartlegge ethvert design på dette ytelsesrommet (figur 1). Design som er dårlige på begge målene blir åpenbart ikke vurdert. I stedet vurderes design som utmerker seg enten i akselerasjon og maksimal hastighet eller i å krysse ulendt terreng - eller som finner en balanse mellom de to. Dette fører til en Pareto-front, et ingeniørbegrep (til ære for den italienske ingeniøren Vilfredo Pareto) definert som settet med design der intet enkelt alternativ kan forbedres i alle aspekter samtidig. Hvis det finnes et kontinuerlig utvalg av kjøretøy langs denne Pareto-fronten, vil sjåfører velge forskjellige modeller basert på deres spesifikke behov.
Jeg baserer mye av den følgende diskusjonen på systembiolog Uri Alons arbeid (Alon 2019; Shoval et al. 2012). La oss forestille oss at vi designer biler: en jeep og en racerbil. De to ytelsesegenskapene vi er interessert i er akselerasjon og evnen til å krysse ulendt terreng. Hvis vi plotter ytelse 1 på x-aksen og ytelse 2 på y-aksen, kan vi kartlegge ethvert design på dette ytelsesrommet (figur 1). Design som er dårlige på begge målene blir åpenbart ikke vurdert. I stedet vurderes design som utmerker seg enten i akselerasjon og maksimal hastighet eller i å krysse ulendt terreng - eller som finner en balanse mellom de to. Dette fører til en Pareto-front, et ingeniørbegrep (til ære for den italienske ingeniøren Vilfredo Pareto) definert som settet med design der intet enkelt alternativ kan forbedres i alle aspekter samtidig. Hvis det finnes et kontinuerlig utvalg av kjøretøy langs denne Pareto-fronten, vil sjåfører velge forskjellige modeller basert på deres spesifikke behov.
Et design eller en fenotype som er maksimalt optimalisert for én egenskap, slik som vi ser i punkt A i figur 1, som er maksimert for ytelse 1 (akselerasjon), kalles en "arketype".
Omvendt ingeniørkunst for å finne optimaliseringskriterier i biologi
På samme måte som i menneskelig ingeniørkunst, finner vi også at biologiske systemer står overfor avveininger. For eksempel er et kjent eksempel på "evolusjon", 17-"arters"-finkestrålingen på Galápagosøyene, et eksempel på en populasjon som beveger seg langs en Pareto-front. Peter og Rosemary Grant samlet inn data fra 1972 til 2001 om "naturlig seleksjon" ved å måle fem variabler av den mellomstore bakkefinken og kaktusfinken på Galápagosøyene (Grant og Rosemary Grant 2002). Disse dataene ble deretter analysert ved hjelp av en omvendt ingeniørkunsttilnærming der finkefenotypene ble kvantifisert i egenskapsrommet. Egenskapsrommet er representert ved å ha en egenskap på x-aksen og en annen på y-aksen, så det ligner på ytelsesrommet.
Slik beskriver Uri Alon denne omvendte konstruksjonen:
"Vi plotter ganske enkelt dataene i egenskaps-rommet, ved å bruke alle egenskapene vi kan måle. Aksene er egenskapene, og hver fenotype er et punkt i dette rommet. For eksempel er hvert nebb et punkt i et rom med egenskaper som nebbbredde, dybde, krumning og så videre." (Alon 2019)
Når dataene er plottet i egenskaps-rommet, vil de anta en form (linje, trekant, polytop), og den formen kan hjelpe oss med å finne ut hvor mange egenskaper som optimaliseres, og til og med hva disse egenskapene kan være. Dette fungerer fordi optimalisering i henhold til oppgaver skaper bestemte geometriske former i egenskapsrommet. Hvis man plotter egenskapsdata og ser en kurve, optimaliseres to oppgaver. Hvis man ser en buet trekant, optimaliseres tre egenskaper. Hvis du ser en buet tetraeder, optimaliseres fire egenskaper. I hjørnene av linjene ligger arketypene eller spesialistene. Design mellom hjørnene er generalister; de er ikke best til å utføre en bestemt oppgave. Likevel kan de 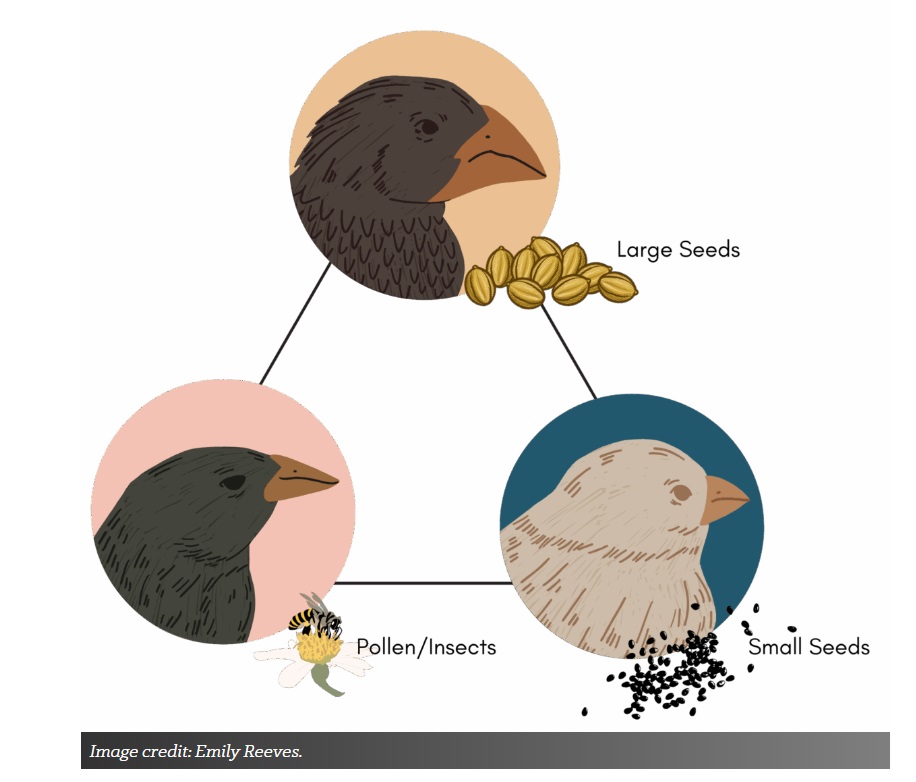 utføre flere oppgaver med en viss dyktighet, noe som også kan være fordelaktig. Ved å se på hva organismene nærmest hjørnene gjør veldig bra, kan man utlede hvilke oppgaver som optimaliseres.
utføre flere oppgaver med en viss dyktighet, noe som også kan være fordelaktig. Ved å se på hva organismene nærmest hjørnene gjør veldig bra, kan man utlede hvilke oppgaver som optimaliseres.
Avveininger for Darwinfinker
På Daphne Major, en Galápagosøy, finnes det tre hovedtyper finkefór: store frø, små frø og pollen og insekter. Noen finker er optimale for å spise bare store frø; disse finkene har et stort nebb og en stor kroppsstørrelse (figur 2). La oss kalle dette arketypen for store frø. Andre finker er optimale for å spise små frø; disse finkene har et lite, kort nebb og en liten kroppsstørrelse. Vi kan kalle dette arketypen for små frø. Andre finker er optimale for å spise pollen og insekter; disse finkene har et rett, spiss nebb og en middels kroppsstørrelse. Og la oss kalle den for pollen/insekt-arketypen. Hver arketype er en spesialist eller den aller beste på en bestemt oppgave. Finken som er optimalisert for å spise store frø er per definisjon ikke optimal for å spise små frø på grunn av avveiningene mellom disse to oppgavene (du kan ikke ha et kroket kort nebb og et langt, tynt nebb samtidig). Finkepopulasjoner kan bevege seg mellom disse arketypene basert på miljøforhold.
Bilde 4. Tre arketyper av finker
Bilde 5. Darwins 'beste' bevis for evolusjon
 Eksisterende genetisk potensial fremfor tilfeldig nyhet
Eksisterende genetisk potensial fremfor tilfeldig nyhet
Etter hvert som arter beveger seg langs pareto-fronten, sier biologer at artene "utvikler seg foran øynene våre". De kommer ofte til den konklusjonen at dette er et eksempel på tilfeldig mutasjon og naturlig seleksjon som skaper nyhet. Likevel, utover det faktum at slik styrt bevegelse motsier hypotesen om tilfeldighet, avslører en nærmere undersøkelse at den genetiske kapasiteten for denne Pareto-banen allerede eksisterer før noe selektivt press. Hvis denne kapasiteten eksisterte før evolusjonen startet, har ingenting virkelig "nyskapende" utviklet seg.
For å forstå det molekylære grunnlaget for finkestrålingen ble genomene til Darwins finker sekvensert på nytt i 2015 av forskere ledet av Peter og Rosemary Grant, sammen med kolleger fra Uppsala universitet (Lamichhaney et al. 2015). Gruppen brukte deretter blandingskartlegging for å avgjøre om finkeegenskapene var arvelige, og i så fall hvilke deler av genomet som var ansvarlige (Rubin et al. 2022). Interessant nok tillot arbeidet deres dem å rapportere at den genetiske variasjonen som ligger til grunn for endringene i finkenebbet, hadde eksistert i populasjonen tidligere:
Bilde 5. Darwins 'beste' bevis på evolusjon
"Vi finner opprinnelsen til disse haplotypeblokkene knyttet til fenotypisk divergenser, forut for artsdannelseshendelser." (Rubin et al. 2022)
Med andre ord oppdaget disse forskerne at haplotypeblokker, DNA-seksjoner ved rekombinasjonskalde punkter, eksisterte før finke- utvidelsen. Fordi blokkene er innenfor rekombinasjonskalde punkter, antas dette å ha tillatt at lociene fortsatte å bevege seg sammen til tross for genflyt mellom populasjoner. Forfatterne bemerker at når den genetiske variasjonen eksisterer i populasjonen tidligere, kan tilpasning skje raskt.
 "Mens rask artsdannelse i adaptiv utstråling gir begrenset tid til å generere de novo genetisk variasjon, kan forfedre-polymorfismer legge til rette for rask akkumulering av ulike kombinasjoner av alleler." (Rubin et al. 2022)
"Mens rask artsdannelse i adaptiv utstråling gir begrenset tid til å generere de novo genetisk variasjon, kan forfedre-polymorfismer legge til rette for rask akkumulering av ulike kombinasjoner av alleler." (Rubin et al. 2022)
Det avgjørende poenget her er at tilfeldig akkumulerende mutasjon ikke førte til strålingen. I stedet ble finketilpasningen som beveget seg langs en paretofront drevet av eksisterende genetisk variasjon.
Bilde 6. Bierer har ofte optimaliserte sosiale algortimer
Trekk og genetikk
De unike finkeegenskapene og genetikken som ligger til grunn for dem er ikke positivt korrelert. I stedet er de negativt korrelert. Dette betyr at allelene som ligger til grunn for tilpasningen er knyttet til avveininger, for eksempel avveiningen mellom å være optimalisert for store frø kontra å være optimalisert for små frø. Sjeldne tilfeldige mutasjoner fører til positive korrelasjoner, men allelene som ligger til grunn for disse egenskapene er knyttet til avveininger eller negative korrelasjoner. Dette støtter ytterligere observasjonen om at genetisk potensial er innebygd i finkepopulasjonen, igjen knyttet til konseptet med avveininger.
Vi ser at i det mest klassiske eksemplet på "evolusjon som skjer foran øynene våre", var genetisk variasjon til stede før den adaptive strålingen. Hvorvidt dette virkelig er "evolusjon" er da et spørsmål verdt å diskutere.
For kilder, se slutten av originalartikkelen -her.
Emily Reeves

Bbilde 7. EMILY REEVES.
Emily Reeves er en biokjemiker, metabolsk ernæringsfysiolog og aspirerende systembiolog. Doktorgradsstudiene hennes ble fullført ved Texas A&M University i biokjemi og biofysikk. Emily er for tiden en aktiv kliniker for metabolsk ernæring og ernæringsgenomikk ved Nutriplexity. Hun liker å identifisere og designe ernæringsintervensjon for subtile medfødte metabolismefeil. Hun jobber også med stipendiater fra Discovery Institute og det større vitenskapelige samfunnet for å fremme integrering av ingeniørvitenskap og biologi. Hun tilbringer helgene sine på tur med mannen sin, brygger kombucha og løper i nærheten av Puget Sound.
Oversettelse, via google oversetter, og bilder ved Asbjørn E. Lund